Receptor T1R2/T1R3: La Dulzura de la Fisiología Humana
Resumen Clínico Rápido
🔬 Clasificación
Receptor Acoplado a Proteína G (GPCR)
⚙️ Función
Percepción del Sabor Dulce
📋 Impacto
Regulación Metabólica y Preferencias Alimentarias
El Receptor de Sabor Dulce T1R2/T1R3: Arquitecto de Nuestra Percepción Glucémica
En el vasto y complejo universo de la fisiología humana, la percepción del sabor dulce emerge como una de las experiencias sensoriales más fundamentales y evolutivamente arraigadas. Esta capacidad, que nos conecta directamente con fuentes de energía vital, no sería posible sin la intrincada maquinaria molecular de los receptores del gusto. Entre ellos, el receptor de sabor dulce T1R2/T1R3 se alza como el principal sensor de azúcares y edulcorantes, una heterodímero de proteínas que orquesta nuestra interacción con la dulzura en el mundo. Su estudio trasciende la mera curiosidad sensorial, adentrándose en profundas implicaciones para la nutrición, el metabolismo y la salud pública, especialmente en contextos dietéticos como la cetosis.
Este receptor, ubicado estratégicamente en las papilas gustativas de la lengua, es mucho más que un simple interruptor de «dulce». Es un sofisticado sistema de señalización que no solo identifica la presencia de glucosa, fructosa y otros carbohidratos, sino que también responde a una miríada de compuestos sintéticos conocidos como edulcorantes artificiales. Comprender su fisiología molecular, su propósito evolutivo y su rol en la modulación de las respuestas metabólicas es crucial para desentrañar los mecanismos subyacentes a nuestras preferencias alimentarias y para desarrollar estrategias dietéticas más efectivas. A través de este análisis profundo, exploraremos cómo este receptor bidimensional moldea nuestra relación con la dulzura y cómo su función se extiende más allá de los límites de la lengua, influenciando sistemas orgánicos clave.
Resumen Clínico
- Punto clave 1: El T1R2/T1R3 es un heterodímero de receptores acoplados a proteína G (GPCR) esencial para la percepción del sabor dulce en humanos.
- Punto clave 2: Se activa por una amplia gama de azúcares (glucosa, fructosa, sacarosa) y edulcorantes artificiales, iniciando una cascada de señalización intracelular.
- Punto clave 3: Más allá de la lengua, se encuentra en tejidos extraorales como el intestino y el páncreas, sugiriendo un rol en la homeostasis glucémica y la regulación metabólica.
Fisiología Molecular del T1R2/T1R3: La Danza de las Proteínas
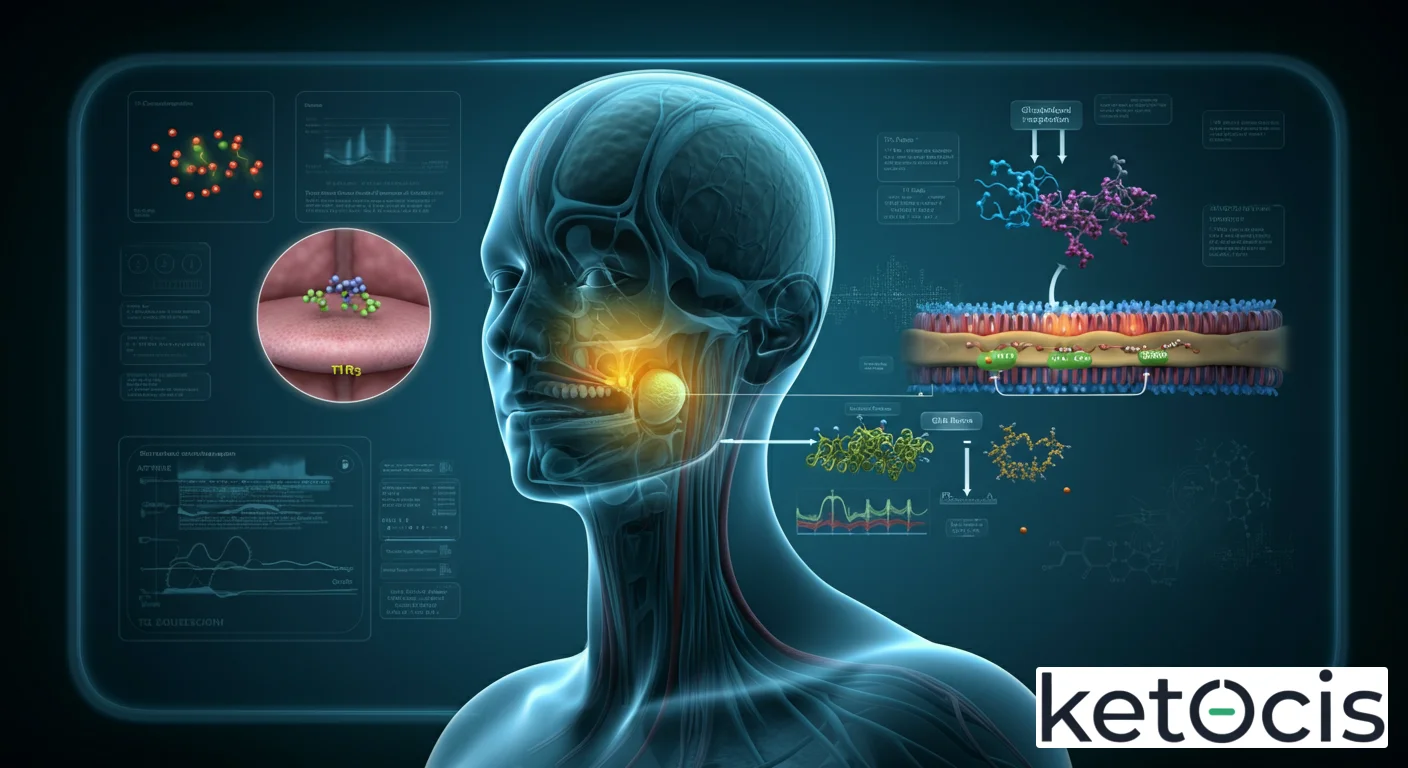
El receptor de sabor dulce T1R2/T1R3 es un ejemplo paradigmático de la complejidad de la quimiorrecepción. Se compone de dos subunidades distintas, la proteína T1R2 y la T1R3, que deben asociarse para formar un receptor funcional. Ambas son miembros de la familia de receptores acoplados a proteína G (GPCRs), específicamente de la Clase C, que incluye receptores de glutamato metabotrópicos y receptores sensibles al calcio. Esta clasificación es importante, ya que dicta su arquitectura molecular y su mecanismo de acción. La subunidad T1R2 es crucial para el reconocimiento de una amplia variedad de edulcorantes orgánicos y artificiales, mientras que la T1R3 confiere la capacidad de responder a azúcares como la glucosa y es indispensable para la funcionalidad del heterodímero.
Cuando un compuesto dulce se une al sitio de unión extracelular del T1R2/T1R3, se induce un cambio conformacional en el receptor. Este cambio activa una proteína G heterotrimérica específica, conocida como gustducina (Gαgust). La gustducina, a su vez, activa la fosfolipasa C beta-2 (PLCβ2), una enzima que hidroliza el fosfatidilinositol 4,5-bisfosfato (PIP2) en diacilglicerol (DAG) e inositol trifosfato (IP3). El IP3 es un segundo mensajero clave que se une a receptores de IP3 en el retículo endoplasmático, provocando la liberación de iones de calcio (Ca2+) almacenados intracelularmente. Este aumento en la concentración de Ca2+ citosólico es el evento central que desencadena la activación del canal de potencial receptor transitorio melastatina 5 (TRPM5).
La activación del canal TRPM5 permite la entrada de sodio (Na+) y, más importantemente, la salida de potasio (K+) y la despolarización de la membrana de la célula gustativa tipo II (célula receptora del sabor dulce). Esta despolarización conduce a la liberación de ATP a través de canales de ATP específicos (como el canal pannexina 1) hacia el espacio extracelular. El ATP liberado actúa como un neurotransmisor, uniéndose a receptores purinérgicos en las fibras nerviosas aferentes del gusto, que son neuronas sensoriales primarias. Estas neuronas transmiten la señal eléctrica al tronco encefálico, luego al tálamo y, finalmente, a la corteza gustativa en el cerebro, donde se interpreta conscientemente como la sensación de dulzura. Este complejo y elegante proceso asegura una detección rápida y precisa de los compuestos dulces, informando al organismo sobre la presencia de nutrientes energéticos.
Los edulcorantes artificiales engañan completamente al cuerpo sin ninguna consecuencia metabólica.
Aunque no aportan calorías, la activación del T1R2/T1R3 por edulcorantes puede inducir respuestas fisiológicas, como la liberación de incretinas o una preparación insulínica, y alterar la microbiota intestinal, afectando la homeostasis glucémica a largo plazo.
El Propósito Evolutivo de la Dulzura: Un Vínculo con la Supervivencia
La capacidad de percibir el sabor dulce no es un mero capricho sensorial; es una adaptación evolutiva profundamente arraigada que ha sido fundamental para la supervivencia de nuestra especie y de muchas otras. En la naturaleza, el sabor dulce es un indicador fiable de la presencia de carbohidratos, las principales fuentes de energía rápida para el metabolismo celular. Las frutas maduras, las mieles y los néctares, ricos en azúcares simples como la glucosa y la fructosa, son intrínsecamente dulces. La atracción innata hacia estos sabores impulsó a nuestros ancestros a buscar y consumir alimentos energéticamente densos, lo que les proporcionaba la energía necesaria para la caza, la recolección, la reproducción y la supervivencia en entornos desafiantes.
El receptor T1R2/T1R3, por lo tanto, no es solo un sensor, sino un guardián de la homeostasis energética. Su activación señaliza al cerebro la llegada inminente de calorías, preparando al cuerpo para su absorción y metabolismo. Esta preferencia por lo dulce se desarrolla temprano en la vida, siendo evidente incluso en los recién nacidos, quienes muestran una clara preferencia por soluciones azucaradas. Este mecanismo asegura que los individuos busquen activamente alimentos ricos en energía, un rasgo vital en épocas de escasez. Sin embargo, en el contexto moderno de abundancia alimentaria y acceso ilimitado a azúcares refinados y edulcorantes artificiales, esta adaptación ancestral puede volverse un arma de doble filo, contribuyendo a la sobrealimentación y a problemas de salud metabólica.
Ubicación y Mecanismos Celulares: Microanatomía de la Percepción
La percepción del sabor dulce se localiza predominantemente en las papilas gustativas de la lengua, estructuras especializadas que contienen conjuntos de células receptoras del gusto. Específicamente, el heterodímero T1R2/T1R3 se expresa en las células gustativas tipo II, también conocidas como células dulces, amargas y umami, que se encuentran dentro de los bulbos gustativos. Estas células son no neuronales, pero son las encargadas de detectar los estímulos químicos y transducirlos en señales eléctricas. Aunque las papilas gustativas están distribuidas por toda la lengua, la densidad de células que expresan T1R2/T1R3 es particularmente alta en la punta de la lengua y en los bordes laterales, lo que contribuye a la topografía de la percepción dulce.
Cada célula gustativa tipo II tiene microvellosidades que se proyectan hacia el poro gustativo, la abertura en la superficie de la lengua que expone los receptores a las sustancias disueltas en la saliva. Es en estas microvellosidades donde se encuentran los receptores T1R2/T1R3. Una vez activado, el receptor inicia la cascada de señalización intracelular previamente descrita, que culmina en la liberación de ATP. Este ATP actúa sobre las fibras nerviosas aferentes del gusto, las cuales pertenecen a los nervios craneales VII (facial), IX (glosofaríngeo) y X (vago), dependiendo de la región de la lengua. La especificidad de la señalización es crucial: mientras que las células tipo II detectan dulce, amargo y umami, las células tipo III detectan ácido y las células tipo I tienen un papel de soporte y, posiblemente, de detección de sal. Esta compartimentación celular asegura una discriminación precisa de los diferentes sabores.
El Receptor Dulce en el Contexto de la Cetosis y el Ayuno
Para aquellos que adoptan dietas cetogénicas o prácticas de ayuno intermitente, la relación con el sabor dulce adquiere una nueva dimensión. En una dieta cetogénica, el consumo de carbohidratos se restringe drásticamente para inducir un estado metabólico de cetosis, donde el cuerpo quema grasas para obtener energía. Esto implica una reducción significativa de azúcares y, por ende, de la activación directa del T1R2/T1R3 por fuentes calóricas dulces. Sin embargo, la percepción y el deseo de dulce persisten, a menudo intensificados al inicio de la adaptación cetogénica, lo que lleva al uso de edulcorantes artificiales.
Los edulcorantes artificiales como la sucralosa, el aspartamo, la sacarina y el eritritol son ampliamente utilizados en dietas bajas en carbohidratos porque activan el receptor T1R2/T1R3, proporcionando la sensación de dulzura sin aportar calorías o impactar directamente la glucemia de manera significativa. Sin embargo, la interacción de estos edulcorantes con el T1R2/T1R3 y sus efectos metabólicos a largo plazo son objeto de intensa investigación y debate. Aunque no elevan la glucosa sanguínea directamente, la activación del receptor dulce puede desencadenar una «respuesta cefálica de insulina», donde el cerebro anticipa la llegada de glucosa y el páncreas libera una pequeña cantidad de insulina. Esta respuesta, si bien generalmente transitoria, podría tener implicaciones para la sensibilidad a la insulina a largo plazo en individuos susceptibles.
Además, la exposición constante a la dulzura, incluso sin calorías, puede mantener activados los circuitos de recompensa del cerebro asociados con el consumo de azúcar, lo que potencialmente perpetúa los antojos de dulce y dificulta la deshabituación a este sabor. En el ayuno, la percepción del dulce puede incluso intensificarse debido a una mayor sensibilidad de los receptores o a cambios en la regulación central del apetito. Comprender cómo el T1R2/T1R3 media estas respuestas es esencial para optimizar las estrategias dietéticas y para educar a los individuos sobre el uso consciente de los edulcorantes en un estilo de vida cetogénico o de ayuno.
Biohacking de la Percepción Dulce
Para aquellos que buscan resetear su paladar y reducir la dependencia del sabor dulce (incluso de edulcorantes), una estrategia efectiva es la «desintoxicación dulce» gradual. Reducir progresivamente la cantidad de edulcorantes y azúcares en la dieta durante varias semanas puede aumentar la sensibilidad del receptor T1R2/T1R3 a la dulzura natural de los alimentos (frutas, verduras). Esto permite disfrutar de sabores más sutiles y reducir la necesidad de estímulos dulces intensos, promoviendo una relación más sana con los alimentos.
Más Allá de la Lengua: Receptores Dulces Ectópicos
Uno de los descubrimientos más fascinantes en la investigación del T1R2/T1R3 es su expresión en tejidos extraorales, es decir, fuera de la cavidad oral. Se ha encontrado que las subunidades T1R2 y T1R3 están presentes en el intestino delgado, el páncreas, el tejido adiposo, el cerebro e incluso en los testículos. Esta distribución ubicua sugiere que el receptor de sabor dulce tiene funciones que van mucho más allá de la mera percepción del gusto, actuando como un sensor metabólico en diversos órganos.
En el intestino, el T1R2/T1R3 se expresa en las células enteroendocrinas L, donde su activación por la presencia de azúcares en la luz intestinal estimula la liberación de hormonas incretinas como el péptido similar al glucagón-1 (GLP-1) y el péptido inhibidor gástrico (GIP). Estas hormonas juegan un papel crucial en la regulación de la glucosa sanguínea, potenciando la secreción de insulina por el páncreas y ralentizando el vaciamiento gástrico. Este mecanismo ilustra cómo el cuerpo detecta la glucosa no solo en la boca, sino también en el tracto digestivo, coordinando una respuesta metabólica temprana y robusta a la ingesta de carbohidratos.
La presencia del T1R2/T1R3 en las células beta pancreáticas también es intrigante. Aunque su función exacta en este contexto aún se está investigando, se postula que podría modular directamente la secreción de insulina en respuesta a la glucosa. En el tejido adiposo, el receptor podría influir en la adipogénesis y el metabolismo de los lípidos. Estos hallazgos abren nuevas vías para comprender la fisiología metabólica y sugieren que la manipulación farmacológica de estos receptores ectópicos podría ofrecer nuevas estrategias para el tratamiento de enfermedades como la diabetes tipo 2 y la obesidad.
Alerta Médica: Edulcorantes Artificiales y Disbiosis Intestinal
Existe una creciente preocupación de que el consumo regular de ciertos edulcorantes artificiales, al activar el T1R2/T1R3 en el intestino, pueda alterar la composición y función de la microbiota intestinal. Estudios sugieren que edulcorantes como la sacarina o la sucralosa pueden inducir disbiosis, impactando negativamente la diversidad bacteriana y favoreciendo especies asociadas con intolerancia a la glucosa y resistencia a la insulina. Aunque no aportan calorías, su interacción con nuestros sistemas biológicos es compleja y podría tener consecuencias metabólicas a largo plazo que aún estamos comprendiendo.
Implicaciones Clínicas y Futuras Direcciones
La comprensión detallada del receptor T1R2/T1R3 tiene profundas implicaciones clínicas. En el ámbito de la nutrición, permite diseñar dietas más informadas, especialmente para personas con diabetes, resistencia a la insulina o aquellos que buscan un control de peso. La elección de edulcorantes, por ejemplo, podría ser más estratégica si se consideran no solo sus propiedades calóricas, sino también sus efectos sobre la microbiota intestinal y las respuestas hormonales mediadas por el T1R2/T1R3 ectópico.
Desde una perspectiva farmacológica, el T1R2/T1R3 representa una diana potencial para el desarrollo de nuevos fármacos. Moduladores alostéricos de este receptor podrían ser utilizados para alterar la percepción del dulce, ya sea potenciándola para reducir el consumo de azúcar o disminuyéndola para mitigar el deseo de dulce. Además, la investigación sobre los receptores T1R2/T1R3 en el intestino está abriendo caminos para desarrollar terapias que optimicen la liberación de incretinas, lo que podría mejorar el control glucémico en pacientes con diabetes. La personalización de la dieta basada en la genética de los receptores gustativos es otra área emergente, ya que existen polimorfismos en los genes T1R2 y T1R3 que pueden influir en la sensibilidad individual al dulce.
Conclusión: La Dulzura como Clave Metabólica
El receptor de sabor dulce T1R2/T1R3 es una maravilla de la ingeniería biológica, un sensor molecular que ha guiado nuestra supervivencia y que continúa influyendo profundamente en nuestra salud y comportamiento alimentario. Desde su función primordial en la lengua, detectando la presencia de azúcares y edulcorantes, hasta su inesperada aparición en órganos metabólicos clave, este heterodímero es un actor central en la compleja red que regula nuestra homeostasis energética. Su estudio nos recuerda que la percepción sensorial no es un fenómeno aislado, sino una parte integral de la fisiología, con ramificaciones que se extienden a cada rincón de nuestro organismo.
A medida que la ciencia continúa desentrañando los misterios del T1R2/T1R3, se abren nuevas avenidas para el biohacking nutricional, la prevención de enfermedades metabólicas y el desarrollo de terapias innovadoras. Comprender y respetar la sofisticación de este receptor es fundamental para navegar el paisaje alimentario moderno y para cultivar una relación más consciente y saludable con uno de los sabores más irresistibles de la naturaleza.
Preguntas Frecuentes Relacionadas
¿Cuánto tiempo tarda el proceso metabólico?
El tiempo varía según el metabolismo individual y la adherencia a la restricción de carbohidratos, pero generalmente toma de 2 a 4 días en condiciones estrictas.
¿Cómo mido mis niveles de forma óptima?
Se recomiendan los medidores de sangre para mayor precisión clínica (miden beta-hidroxibutirato), aunque existen opciones de aliento y tiras de orina para principiantes.
¿Es normal sentir fatiga al inicio?
Sí, durante la fase de adaptación es común experimentar la «gripe keto». Mantener una óptima hidratación y reponer electrolitos (sodio, potasio, magnesio) mitiga drásticamente estos efectos.
Explorar Glosario Médico
Explora Nuestros Centros Temáticos
Tu Panel Metabólico
Sincroniza tus registros y monitorea tu progreso en tiempo real con Ketocis Tracker.
Directorio de Recetas Keto
Explora nuestra base de datos completa de platillos bajos en carbohidratos.