¿Qué es el factor de crecimiento similar a la insulina 1 (IGF-1) local? – Análisis Completo y Beneficios
Resumen Clínico Rápido
🔬 Clasificación
Polipéptido, Factor de Crecimiento
⚙️ Función
Crecimiento, Reparación Tisular, Supervivencia Celular
📋 Impacto
Regulación Autocrina/Paracrina, Salud Tisular
¿Qué es el Factor de Crecimiento Similar a la Insulina 1 (IGF-1) Local?
En el complejo y dinámico universo de la fisiología humana, el Factor de Crecimiento similar a la Insulina 1 (IGF-1) emerge como una molécula de inmensa importancia. Sin embargo, su comprensión a menudo se simplifica, centrándose exclusivamente en su forma sistémica, aquella producida principalmente por el hígado bajo la influencia de la hormona del crecimiento (GH). Esta perspectiva, aunque fundamental, omite una faceta crítica y fascinante: el IGF-1 local o tisular. Esta guía enciclopédica desentrañará la naturaleza, función y relevancia del IGF-1 local, proporcionando una visión profunda y matizada esencial para cualquier investigador, clínico o entusiasta del biohacking.
El IGF-1 es un polipéptido con una estructura molecular homóloga a la insulina, lo que le confiere su nombre. Actúa como un potente mediador del crecimiento y desarrollo, influyendo en la proliferación, diferenciación y supervivencia celular en prácticamente todos los tejidos. Mientras que el IGF-1 circulante, de origen hepático, modula el crecimiento esquelético y metabólico general, el IGF-1 local opera con una autonomía relativa, orquestando respuestas específicas en el microambiente tisular. Comprender esta distinción es crucial para diseñar estrategias de intervención precisas que busquen optimizar la salud y longevidad, especialmente en contextos metabólicos como la cetosis y el ayuno.
La investigación moderna subraya que el IGF-1 no es una entidad monolítica. Su acción está finamente regulada por su origen, sus proteínas de unión (IGFBPs) y la densidad de sus receptores (IGF-1R) en las células diana. La dicotomía entre el IGF-1 endocrino (sistémico) y el IGF-1 autocrino/paracrino (local) es una piedra angular para entender cómo el cuerpo mantiene el equilibrio entre el crecimiento global y la reparación tisular específica. En el Glosario Ketocis, esta diferenciación es vital, ya que las dietas cetogénicas y los períodos de ayuno pueden modular de forma distinta ambos compartimentos de IGF-1, con implicaciones profundas para la salud metabólica y la función celular.
Resumen Clínico
- El IGF-1 local es producido directamente por los tejidos y actúa de forma autocrina o paracrina, diferenciándose del IGF-1 sistémico de origen hepático.
- Su función principal es la reparación tisular, el crecimiento celular específico y la supervivencia en el microambiente local, a menudo con independencia parcial de la hormona del crecimiento.
- La modulación del IGF-1 local mediante ejercicio y nutrientes específicos es clave para optimizar la regeneración muscular y ósea, incluso en estados de ayuno o cetosis.
El IGF-1 es siempre 'malo' o 'pro-cáncer' y debe ser suprimido por completo para la longevidad.
El IGF-1 sistémico elevado crónicamente se asocia con riesgos, pero el IGF-1 local es esencial para la regeneración y reparación de tejidos como el músculo, hueso y cerebro. Su supresión total sería perjudicial para la salud y función tisular. La clave es el equilibrio y la especificidad de acción.
Origen del IGF-1 Local: Un Orquestador Tisular
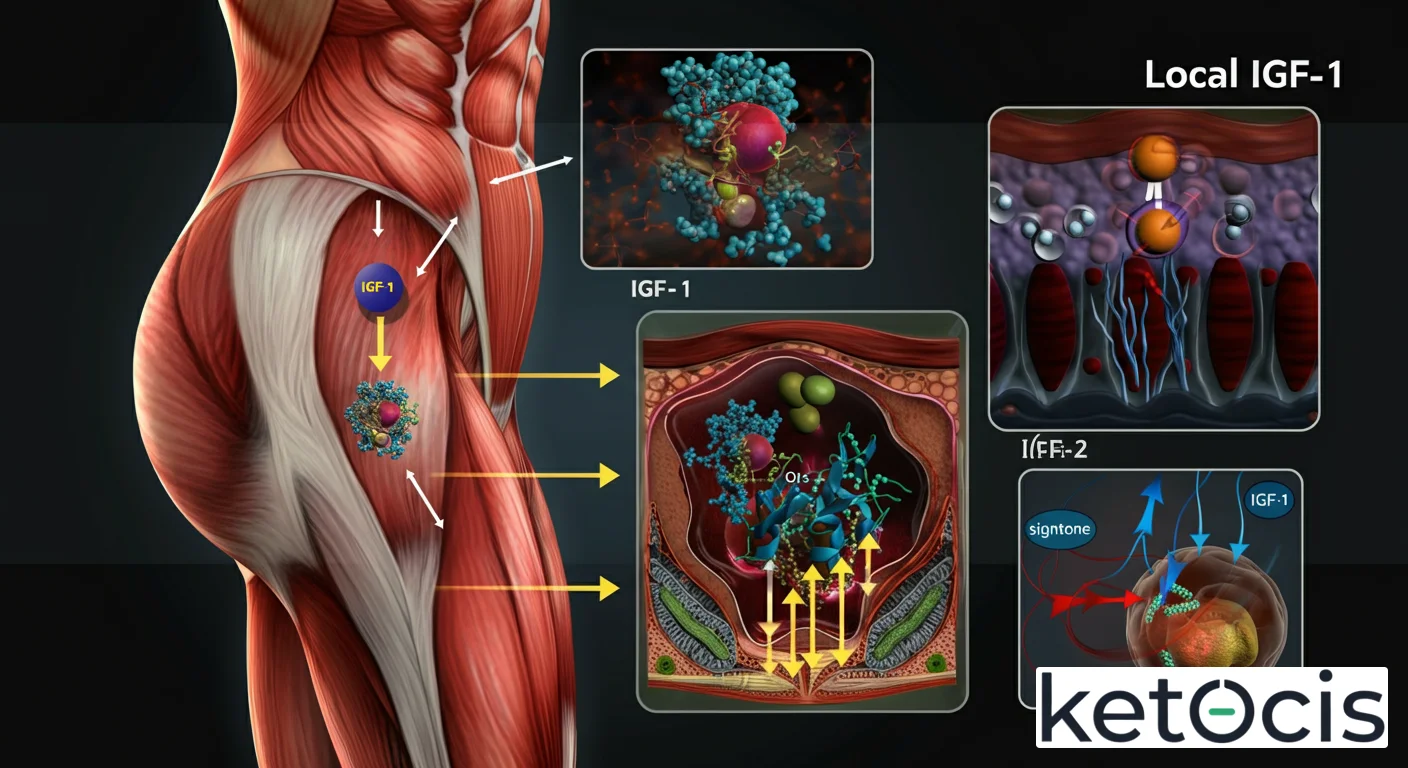
A diferencia del IGF-1 sistémico, cuya síntesis está predominantemente anclada al hígado y fuertemente regulada por la hormona del crecimiento (GH), el IGF-1 local se produce en una vasta red de tejidos a lo largo del cuerpo. Cada célula, en cierta medida, tiene la capacidad de sintetizar su propio IGF-1 en respuesta a estímulos específicos de su entorno. Esto incluye, pero no se limita a, el músculo esquelético, el hueso, el cartílago, el cerebro, el corazón, los riñones, los pulmones, la piel y el sistema inmune.
La producción de IGF-1 local es un proceso altamente regulado y adaptativo. Por ejemplo, en el músculo esquelético, el estrés mecánico inducido por el ejercicio de resistencia es un potente estímulo para la expresión y secreción de IGF-1 de tipo MGF (factor de crecimiento mecánico), una isoforma específica que juega un papel crucial en la reparación y el crecimiento de las fibras musculares. Este IGF-1 de origen muscular actúa localmente para promover la proliferación y diferenciación de las células satélite, esenciales para la hipertrofia y la regeneración.
En el tejido óseo, los osteoblastos (células formadoras de hueso) y los osteocitos (células óseas maduras) producen IGF-1 en respuesta a la carga mecánica y a otros factores de crecimiento. Este IGF-1 autóctono es fundamental para mantener la densidad ósea y para los procesos de remodelación. De manera similar, en el cerebro, neuronas y células gliales sintetizan IGF-1, que es vital para la neurogénesis, la plasticidad sináptica y la protección neuronal, actuando como un factor neurotrófico.
Es importante destacar que la síntesis de IGF-1 local puede ser modulada por factores que no afectan necesariamente al IGF-1 sistémico. Por ejemplo, la inflamación local, la disponibilidad de nutrientes específicos, la presencia de otros factores de crecimiento locales (como PDGF, FGF) y las citoquinas pueden influir en su producción. Esta autonomía parcial confiere al IGF-1 local una capacidad única para mediar respuestas adaptativas y de reparación en el contexto específico de cada tejido, lo que lo convierte en un objetivo terapéutico y de biohacking de gran interés.
Mecanismo de Acción: Señalización Precisa en el Microambiente
El IGF-1 local ejerce sus efectos biológicos principalmente uniéndose a su receptor de alta afinidad, el receptor IGF-1R, una tirosina quinasa transmembrana. Aunque el IGF-1R tiene una estructura similar al receptor de insulina y puede interactuar con él, las vías de señalización que activa son distintivas y cruciales para sus funciones específicas.
Una vez que el IGF-1 se une al IGF-1R, se produce una autofosforilación de los residuos de tirosina en el dominio intracelular del receptor. Esto recluta proteínas adaptadoras, como los sustratos del receptor de insulina (IRS) y las proteínas Shc, que a su vez inician cascadas de señalización intracelular. Las dos vías principales activadas por el IGF-1R son la vía de la fosfatidilinositol 3-quinasa (PI3K)/Akt/mTOR y la vía de las proteínas quinasas activadas por mitógenos (MAPK/ERK).
La vía PI3K/Akt/mTOR es fundamental para promover el crecimiento celular, la síntesis de proteínas, la supervivencia celular y la inhibición de la apoptosis. La activación de Akt, una serina/treonina quinasa, conduce a la fosforilación de múltiples sustratos que regulan el metabolismo de la glucosa, la síntesis de lípidos y, crucialmente, la activación de mTOR (objetivo de rapamicina en mamíferos). mTOR es un complejo central en la regulación del crecimiento celular y la homeostasis proteica, y su modulación por el IGF-1 local es esencial para la regeneración tisular y la hipertrofia.
Por otro lado, la vía MAPK/ERK está más implicada en la proliferación y diferenciación celular. La activación de estas quinasas conduce a la expresión de genes que regulan el ciclo celular y procesos de desarrollo específicos del tejido. La acción combinada y equilibrada de estas dos vías permite que el IGF-1 local orchestre respuestas celulares complejas y altamente específicas, adaptadas a las necesidades del microambiente tisular.
Además de la señalización directa a través del IGF-1R, la biodisponibilidad del IGF-1 local está intrínsecamente regulada por las proteínas de unión al IGF (IGFBPs). Existen seis IGFBPs principales (IGFBP-1 a -6), que se unen al IGF-1 con alta afinidad. Estas proteínas no solo transportan el IGF-1 y prolongan su vida media, sino que también modulan su acceso al receptor, pudiendo potenciar o inhibir sus acciones. La expresión de IGFBPs es también local y puede variar según el tejido y el estado fisiológico, añadiendo otra capa de complejidad a la regulación del IGF-1 local.
Antagonistas y Reguladores del IGF-1 Local
La actividad del IGF-1 local no es un proceso descontrolado; está sujeta a una compleja red de antagonistas y mecanismos reguladores que garantizan una homeostasis tisular precisa. Entender estos reguladores es tan importante como comprender el propio factor de crecimiento para cualquier estrategia de optimización.
Como se mencionó, las proteínas de unión al IGF (IGFBPs) son los principales reguladores de la biodisponibilidad del IGF-1. Algunas IGFBPs, como la IGFBP-1 y la IGFBP-2, generalmente inhiben la acción del IGF-1 al secuestrarlo y prevenir su unión al receptor. Otras, como la IGFBP-3, que es la más abundante en el suero, pueden tener efectos duales: prolongar la vida media del IGF-1 y, en ciertos contextos, facilitar su entrega a las células. La expresión de estas IGFBPs es regulada localmente por factores como la disponibilidad de nutrientes, hormonas y citoquinas, permitiendo un control fino sobre la acción del IGF-1 en cada tejido.
Hormonas sistémicas también influyen, aunque indirectamente, en el IGF-1 local. Aunque el IGF-1 local puede operar con cierta independencia de la GH, niveles crónicamente bajos o altos de hormona del crecimiento pueden alterar el entorno metabólico general y, en consecuencia, la capacidad de los tejidos para producir o responder al IGF-1. La insulina, por ejemplo, es un potente regulador de IGFBP-1 en el hígado, y sus niveles influirán en la disponibilidad de IGF-1 circulante, lo que a su vez puede tener efectos indirectos en la regulación local.
Los glucocorticoides, como el cortisol, son conocidos por sus efectos catabólicos y pueden antagonizar la acción del IGF-1 al inhibir su síntesis y promover la degradación proteica. En situaciones de estrés crónico o inflamación, los niveles elevados de cortisol pueden suprimir la señalización del IGF-1, lo que afecta negativamente la reparación tisular y la función inmune. Por otro lado, la inflamación aguda puede modular positivamente el IGF-1 local en el sitio de la lesión para facilitar la curación.
A nivel celular, las vías de señalización que activan el IGF-1 son también sujetas a regulación negativa. Por ejemplo, la proteína PTEN (phosphatase and tensin homolog) antagoniza la vía PI3K/Akt al desfosforilar PIP3, un lípido esencial para la activación de Akt. Esto limita la señalización proliferativa y de supervivencia, actuando como un supresor tumoral y un regulador de la sensibilidad a la insulina. La actividad de PTEN es crucial para prevenir el crecimiento celular descontrolado.
Finalmente, los sensores de nutrientes celulares, como mTOR y AMPK, juegan un papel bidireccional. Mientras que el IGF-1 local activa mTOR para promover el crecimiento, la AMPK, activada por la escasez de energía (como en el ayuno o el ejercicio intenso), inhibe mTOR y otras vías anabólicas, lo que puede modular la respuesta al IGF-1. Este delicado equilibrio entre las vías anabólicas (IGF-1/mTOR) y catabólicas/recicladoras (AMPK/autofagia) es fundamental para la homeostasis energética y la longevidad.
Biohacking y Optimización del IGF-1 Local en el Contexto de Ketocis
La distinción entre el IGF-1 sistémico y el IGF-1 local es particularmente relevante para la comunidad de biohacking y aquellos interesados en la cetosis y el ayuno. Mientras que la reducción del IGF-1 sistémico se asocia a menudo con beneficios de longevidad y una menor incidencia de ciertas enfermedades (mediado por la inhibición de mTOR), el IGF-1 local es indispensable para la reparación tisular, la función muscular y la salud cerebral.
El objetivo del biohacking no es eliminar el IGF-1, sino modular su expresión y acción de manera inteligente. La estrategia es maximizar los beneficios del IGF-1 local para la salud tisular, la recuperación y el rendimiento, mientras se mantiene bajo control el IGF-1 sistémico, si es ese el objetivo para la longevidad.
Estrategias de Optimización:
-
Entrenamiento de Resistencia Localizado: El ejercicio de fuerza es el estímulo más potente conocido para la producción de IGF-1 local en el músculo esquelético. Al generar estrés mecánico y microlesiones, se activa la expresión de MGF (Mechano Growth Factor), una isoforma de IGF-1 que actúa específicamente en la reparación y el crecimiento muscular. La clave es la especificidad: los músculos entrenados producirán más IGF-1 local en respuesta al estímulo.
-
Ingesta de Proteínas y Aminoácidos Esenciales: Aunque el ayuno y la cetosis pueden reducir el IGF-1 sistémico, una ingesta adecuada y estratégica de proteínas, especialmente con un alto contenido de leucina (un potente activador de mTOR), es crucial para apoyar la síntesis de proteínas musculares y la respuesta al IGF-1 local post-ejercicio. Consumir proteínas de calidad en la ventana anabólica post-entrenamiento puede optimizar la respuesta sin comprometer los beneficios de un IGF-1 sistémico bajo.
-
Nutrientes Específicos y Micronutrientes: Ciertos micronutrientes, como el zinc, el magnesio y las vitaminas D y B6, son cofactores importantes en la síntesis de IGF-1 y en las vías de señalización anabólicas. Optimizar su ingesta puede apoyar la función del IGF-1 local. Los ácidos grasos omega-3 también pueden influir positivamente en la sensibilidad al IGF-1 y reducir la inflamación, lo que indirectamente favorece la reparación tisular.
-
Optimización del Sueño y Reducción del Estrés: El sueño de calidad es fundamental para la recuperación y la producción de GH, que, aunque más relacionada con el IGF-1 sistémico, también influye en la capacidad general de los tejidos para repararse. El estrés crónico y los niveles elevados de cortisol pueden suprimir la acción del IGF-1, por lo que las estrategias de manejo del estrés (meditación, mindfulness) son indirectamente beneficiosas.
-
Ciclos de Nutrición (Targeted Ketogenic Diet, Ayuno Intermitente): Para aquellos que buscan los beneficios de la cetosis y el ayuno para la longevidad (reducción de mTOR y IGF-1 sistémico), la integración de «re-feeds» estratégicos o ciclos de carbohidratos/proteínas en momentos específicos (por ejemplo, después del entrenamiento) puede permitir una elevación transitoria del IGF-1 y la insulina para impulsar la recuperación y el crecimiento muscular, sin mantener elevados los niveles sistémicos de forma crónica.
Es fundamental recordar que la modulación del IGF-1 es un acto de equilibrio. El exceso crónico de IGF-1, especialmente el sistémico, se ha asociado con un mayor riesgo de ciertas patologías. Por ello, el biohacking debe ser siempre informado y, preferiblemente, guiado por un profesional de la salud.
Conclusión: El IGF-1 Local como Pilar de la Salud Celular
El Factor de Crecimiento similar a la Insulina 1 (IGF-1) local representa una dimensión fundamental de la fisiología humana que a menudo se subestima en favor de su contraparte sistémica. Hemos explorado cómo este polipéptido, producido autóctonamente por una miríada de tejidos, actúa como un orquestador maestro de la reparación, el crecimiento y la supervivencia celular en el microambiente específico. Su capacidad para operar con una autonomía relativa de la hormona del crecimiento y la insulina sistémica le confiere un papel único en la adaptación y resiliencia de nuestro organismo.
Desde la regeneración muscular post-ejercicio hasta la neuroprotección y el mantenimiento de la densidad ósea, el IGF-1 local es un pilar indispensable para la salud tisular. Su mecanismo de acción, mediado por la activación de vías de señalización clave como PI3K/Akt/mTOR y MAPK/ERK, permite una respuesta celular precisa y adaptada a las demandas locales. Además, la compleja red de proteínas de unión (IGFBPs) y otros reguladores hormonales y metabólicos asegura que la actividad del IGF-1 local esté finamente sintonizada, evitando tanto la deficiencia como el exceso descontrolado.
Para la comunidad de Glosario Ketocis y los practicantes del biohacking, comprender la dualidad del IGF-1 (sistémico vs. local) es empoderador. Permite diseñar estrategias inteligentes que pueden apuntar a los beneficios de la longevidad asociados con un IGF-1 sistémico más bajo (típico en cetosis y ayuno), mientras se optimiza simultáneamente el IGF-1 local para mantener la integridad, función y capacidad de reparación de tejidos vitales. El entrenamiento de resistencia, una nutrición proteica estratégica y la atención a micronutrientes y factores de estilo de vida como el sueño emergen como herramientas poderosas para manipular este equilibrio.
En última instancia, el IGF-1 local no es simplemente una molécula de crecimiento; es un testimonio de la intrincada sabiduría del cuerpo para mantener la homeostasis y la capacidad de regeneración. Su estudio y comprensión profundos abren nuevas avenidas para la medicina preventiva, las terapias regenerativas y la optimización de la salud a lo largo de la vida, cimentando su lugar como uno de los factores más fascinantes y vitales en el panorama de la biología humana.
Preguntas Frecuentes Relacionadas
¿Cuánto tiempo tarda el proceso metabólico?
El tiempo varía según el metabolismo individual y la adherencia a la restricción de carbohidratos, pero generalmente toma de 2 a 4 días en condiciones estrictas.
¿Cómo mido mis niveles de forma óptima?
Se recomiendan los medidores de sangre para mayor precisión clínica (miden beta-hidroxibutirato), aunque existen opciones de aliento y tiras de orina para principiantes.
¿Es normal sentir fatiga al inicio?
Sí, durante la fase de adaptación es común experimentar la «gripe keto». Mantener una óptima hidratación y reponer electrolitos (sodio, potasio, magnesio) mitiga drásticamente estos efectos.
Explorar Glosario Médico
Explora Nuestros Centros Temáticos
Tu Panel Metabólico
Sincroniza tus registros y monitorea tu progreso en tiempo real con Ketocis Tracker.
Directorio de Recetas Keto
Explora nuestra base de datos completa de platillos bajos en carbohidratos.